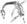Chainosauria
Chainosauria

Crâne fossile de Galepus (en).
| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Classe | Synapsida |
| Ordre | Therapsida |
| Sous-ordre | † Anomodontia |
Clade
Taxons de rang inférieur
Liste
- † Galepus (en)
- † Galeops (en)
- † Galechirus
- † Patranomodon ?
- † Anomocephaloidea ?
- † Dicynodontia
Les chainosaures, ou chainosauriens (Chainosauria), forment un clade éteint de thérapsides anomodontes qui incluent les dicynodontes et certains genres basaux apparentées. Le taxon fut initialement érigée en 1923 par Franz Nopcsa afin de regrouper les dicynodontes et leur proches parents, à savoir trois genres de petits anomodontes connues d'Afrique du Sud (Galepus (en), Galeops (en) et Galechirus) qui constituait le groupe aujourd'hui obsolète des « Dromasauria ». Cependant, ce nom tomba rapidement en désuétude, étant remplacé fonctionnellement par Anomodontia. Le taxon Chainosauria est ensuite relancé cladistiquement à partir de 2009, préservant l'association des dicynodontes et des « dromasauriens » et a depuis servi en effet à la fois de contrepartie cladistique et biogéographique aux venyukovioïdes laurasiens, les premiers chainosaures semblant avoir eu une diversification d'origine gondwanienne.
D'autres lignées d'anomodontes basaux, à savoir le groupe des anomocéphaloïdes et le genre Patranomodon, ont parfois été également récupérés en tant que membre de ce clade, de sorte que les relations des premiers chainosauriens soient irrésolus. En tant que telles, les autapomorphies (traits déterminants uniques) qui unissent les chainosaures sont difficiles à déterminer avec certitude. Les premiers chainosaures sont beaucoup plus légers et élancés que les dicynodontes. Néanmoins, leurs crânes ressemblent généralement et proportionnellement plus à ceux des dicynodontes qu'aux anomodontes plus basaux. Les premiers chainosauriens ne possèdent pas les défenses et le bec caractéristiques des dicynodontes, se disposants même à l'inverse des rangées complètes de dents. Cependant, l'articulation coulissante de la mâchoire et la musculature de cette dernière chez des dicynodontes évolua d'abord chez les chainosauriens antérieurs (à savoir chez l'adjacent Galeops), réduisant et simplifiant leurs dents (en dehors des anomocéphaloïdes).
Historique des recherches

Le taxon Chainosauria est érigée par le paléontologue austro-hongrois Franz Nopcsa en 1923 comme un ordre au sein des Theromorpha (un groupe à peu près équivalent de Therapsida) pour inclure les dicynodontes et les « dromasauriens ». Dans sa classification, Nopcsa inclut également à l'origine le non apparenté sphenacodonte Palaeohatteria, le plaçant dans les « Dromasauria » au sein de sa propre famille des Palaeohatteriidae, tandis que les autres « dromasaures » forment les Dromasauridae[1],[2].
Le concept proposée par Nopcsa pour les Chainosauria englobant les dicynodontes et leurs proches parents était fonctionnellement similaire à l'usage moderne d'Anomodontia, qui était lui-même utilisé à l'époque de manière équivalente à ce qui est actuellement reconnu sous le nom de Dicynodontia. En tant que tel, la redéfinition et l'expansion ultérieure d'Anomodontia par d'autres chercheurs pour englober les dicynodontes et leurs proches parents ont rendu Chainosauria redondant, et le nom est donc tombé en désuétude pour le reste du XXe siècle[3]. La validité de Chainosauria fut même critiquée par le paléontologue américain Alfred Romer la même année durant lequel le taxon fut érigée, considérant le choix de Nopcsa de regrouper les dicynodontes et les « dromasauriens » comme étant « pour des raisons peu évidentes »[4]. Cependant, une véritable relation phylogénétique entre les « dromasaures » et les dicynodontes a depuis été confirmée[5]. De plus, à mesure que davantage d'anomodontes basaux ont été découverts et que de nouvelles méthodes cladistiques pour déterminer les relations phylogénétiques ont été développées, une association étroite entre les « dromasaures » et les dicynodontes, à l'exclusion des autres anomodontes, a également été reconnue[6].
Cela a conduit les paléontologues Christian F. Kammerer et Kenneth D. Angielczyk à ressusciter le nom en 2009 pour ce groupe phylogénétique. Ils redéfinissent cladistiquement Chainosauria comme le clade de tous les anomodontes plus proche de Dicynodon et Galeops (en) que de Venyukovia (en), définissant en fait Chainosauria comme un homologue du clade d'anomodontes russe des Venyukovioidea. Tel que conçu par Kammerer et Angielczyk, Chainosauria excluait également les anomodontes basaux sud-africains Anomocephalus et Patranomodon. Cependant, des analyses phylogénétiques récentes ont permis de retrouver Patranomodon ainsi que les anomocéphaloïdes comme plus proches des dicynodontes que des venyukovioïdes et donc comme des chainosaures par définition[7],[8],[9].
Morphologie
La majorité de la diversité des chainosauriens se trouve chez les dicynodontes, qui couvrent une large gamme de tailles corporelles mais sont de forme conservatrice, généralement avec un corps en forme de tonneau, des membres trapus et une queue courte. En revanche, les chainosaures antérieurs sont beaucoup plus légers et graciles, avec des os des membres allongés et minces[10]. De longues queues sont également connues au moins chez Galepus (en) et Galechirus, qui ont chacun environ 30 vertèbres caudales qui sont plus longues que larges (contrairement aux vertèbres caudales courtes et larges des dicynodontes, mais similaires à celles du venyukovioïde Suminia)[5],[11].
Les premiers chainosaures n'avaient pas encore développé la dentition postcanine réduite, les becs et les défenses des dicynodontes, et possédaient à la place des rangées complètes de dents dépourvues de canines différenciées (à l'exception potentielle de l'anomocéphaloïde Tiarajudens). La dentition des premiers chainosaures définitifs (c'est-à-dire les « dromasauriens »), ainsi que de Patranomodon, est constituée de dents simplifiées en forme de chevilles[5]. Une exception possible sont les anomocéphaloïdes, qui possèdent des dents incisiformes en forme de feuille et des dents palatines en forme de molaire (ainsi que des dents caniniformes en forme de sabre chez les Tiarajudens), s'ils sont effectivement des chainosaures. Notamment, cependant, l'articulation allongée caractéristique de la mâchoire inférieure des dicynodontes et la nouvelle musculature de la mâchoire associée (l'adducteur latéral externe) qui permet à la mandibule de glisser vers l'arrière et de traiter les aliments se trouvent pour la première fois à l'extérieur de Dicynodontia chez Galeops. Un bec naissant au bout des mâchoires a parfois été proposé chez Galeops et les Galepus (en), mais il s'agit probablement d'une partie anatomique mal conservé[12].
Le nerf naso-palatin à l'intérieur du maxillaire est déjà volumineux et bien développé chez Patranomodon. Ce nerf est une branche du nerf trijumeau et est associé à l'innervation du bec kératineux chez les dicynodontes (semblables aux tortues), et son élargissement chez Patranomodon peut indiquer qu'une plus grande sensibilité dans le prémaxillaire aurait pu précéder le développement des becs chez les premiers chainosauriens[13].
Classification
Phylogénétique
Bien que définis comme incluant les dicynodontes et les « dromasauriens », les « dromasauriens » eux-mêmes sont une série (ou un grade) paraphylétique ou même une regroupement polyphylétique de premiers chainosaures en dehors des dicynodontes[14],[3]. Dans une définition cladistique, Chainosauria comprend tous les anomodontes plus étroitement liés à Dicynodon (représentant les dicynodontes) et à Galeops (en) (représentant les « dromasauriens ») qu'à Venyukovia (en), et est défini de telle sorte que le groupe soit toujours vue en tant que taxon frère des Venyukovioidea. Cela signifie que Chainosauria comprendrait également d'autres anomodontes non considérés comme des dicynodontes ou des « dromasauriens » tant qu'ils se révèlent plus proches d'eux que des venyukovioïdes[3].

En raison des relations instables entre les anomodontes basaux, cela a conduit à des compositions variables pour Chainosauria. L'anomodonte basal Patranomodon fut particulièrement instable à cet égard. Il fut d'abord admis qu'il s'agissait de l'anomodonte le plus basal lors de sa découverte, une interprétation étayée par les premières analyses phylogénétiques. Cependant, plusieurs analyses plus récentes publiées à partir de 2009 le trouvent plutôt comme un chainosaure niché parmi divers « dromasauriens »[11],[7]. De même, Anomocephalus était considéré comme un anomodonte basal en dehors du clade réunissant les Chainosauria et les Venyukovioidea, mais à la suite d'une analyse effectuée en 2017, ce dernier et son proche parent Tiarajudens (formant tout deux les Anomocephaloidea) ont également été récupérés en tant que chainosauriens basaux[15],[16].
L'inclusion de Patranomodon et des anomocéphaloïdes dans les Chainosauria est décrite par Angielczyk et ses collègues en 2017 comme « intuitivement agréable ». Les deux taxons ont des proportions crâniennes profondes et courtes similaires à celles des autres chainosauriens, et bon nombre de leurs caractéristiques basales supposées pourraient également être interprétées comme des plésiomorphies, c'est-à-dire des traits ancestraux partagés et hérités de l'ancêtre commun des chainosaures et d'autres anomodontes. Dans le même temps, les traits dérivés partagés entre les chainosaures et les veinyukovioïdes dérivés (à savoir ceux de l'articulation et la musculature de la mâchoire) sont probablement issues d'une convergence évolutive[16].
Les relations entre les différents « dromasauriens » sont également perturbées. Galeops est systématiquement reconnu comme le taxon frère de Dicynodontia, mais Galepus (en) et Galechirus sont trouvés dans différentes positions. Initialement, ils furent récupérés comme étant étroitement liés à Galeops et aux dicynodontes, comme dans une étude menée par Kammerer et ses collègues en 2011, et formant parfois un clade ensemble sous le nom de Galechiridae[6]. Cependant, dans des analyses ultérieures, Galechirus avait tendance à se situer complètement en dehors des Chainosauria dans une polytomie avec lui et Venyukovioidea (comme dans le cladogramme de gauche ci-dessous). De plus, dans une analyse plus récente de 2017 avec une définition élargie des Chainosauria, Galechirus revient à une position proche de Galeops, tandis que Galepus fut plutôt trouvé comme le chainosaurien le plus basal[16].
Des exemples de ces relations variables sont présentés dans les deux cladogrammes ci-dessous :
| Cladogramme montrant une définition restreinte des Chainosauria selon Cisneros et al., (2015)[15] :
| Cladogramme montrant une définition élargie des Chainosauria selon Angielczyk & Kammerer (2017)[16] :
|
Biogéographie
La composition de Chainosauria a des implications sur les origines, l'histoire évolutive et la biogéographie des premiers anomodontes. Une division biogéographique précoce entre les anomodontes basaux fut proposée par les chercheurs entre les Chainosauria et les Venyukovioidea, les chainosauriens se diversifiant dans le Gondwana dans l'hémisphère sud et les venyukovioïdes se diversifiant en Laurasie dans l'hémisphère nord[7]. Patranamodon et Anomocephalus, ayant chacun été parfois considérés comme les anomodontes les plus basaux, furent utilisés pour argumenter sur une origine gondwanienne des chainosaures et des venyukovioïdes[14],[17]. La reconnaissance du taxon chinois Biseridens en tant qu'anomodonte le plus basal connu soutient alternativement une origine laurasienne pour les anomodontes, d'où les chainosaures auraient migré hors du Gondwana et n'y auraient prospéré qu’ultérieurement[7].
Notes et références
Notes
(en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Chainosauria » (voir la liste des auteurs).
Références
- ↑ (de) Franz Nopcsa, « Die Familien der Reptilien », Fortschritte der Geologie und Palaeontologie, vol. 2, , p. 1-210 (lire en ligne).
- ↑ (en) Frederik Spindler, « Morphological description and taxonomic status of Palaeohatteria and Pantelosaurus (Synapsida: Sphenacodontia) », Freiberger Forschungshefte, vol. C550, no 23, , p. 1-57 (lire en ligne).
- ↑ a b et c (en) Christian F. Kammerer et Kenneth D. Angielczyk, « A proposed higher taxonomy of anomodont therapsids », Zootaxa, vol. 2018, , p. 1-24 (DOI 10.11646/ZOOTAXA.2018.1.1, S2CID 55450468, lire en ligne [PDF]).
- ↑ (en) Alfred S. Romer, « Die Familien der Reptilien. Franz Baron Nopcsa », The Journal of Geology, vol. 31, no 9, , p. 688-689 (DOI 10.1086/623061, S2CID 132493016).
- ↑ a b et c (en) Donald Brinkman, « The structure and relationships of the dromasaurs (Reptila: Therapsida) », Breviora, vol. 465, , p. 1-34 (lire en ligne).
- ↑ a et b (en) Christian F. Kammerer et Kenneth D. Angielczyk, « A comprehensive taxonomic revision of Dicynodon (Therapsida, Anomodontia) and its implications for dicynodont phylogeny, biogeography, and biostratigraphy », Journal of Vertebrate Paleontology, vol. 31, no Suppl. 1, , p. 1-158 (DOI 10.1080/02724634.2011.627074, S2CID 84987497).
- ↑ a b c et d (en) Jun Liu, Bruce Rubidge et Jinling Li, « A new specimen of Biseridens qilianicus indicates its phylogenetic position as the most basal anomodont », Proceedings of the Royal Society B, vol. 277, no 1679, , p. 285-292 (PMID 19640887, PMCID 2842672, DOI 10.1098/rspb.2009.0883).
- ↑ (en) Kenneth D. Angielczyk et Bruce S. Rubidge, « A new pylaecephalid dicynodont (Therapsida, Anomodontia) from the Tapinocephalus Assemblage Zone, Karoo Basin, Middle Permian of South Africa », Journal of Vertebrate Paleontology, vol. 30, no 5, , p. 1396-1409 (DOI 10.1080/02724634.2010.501447, S2CID 129846697).
- ↑ (en) Juan C. Cisneros, Fernando Abdala, Bruce S. Rubidge, Paula C. Dentzien-Dias et Ana de Oliveira Bueno, « Dental occlusion in a 260-million-year-old therapsid with saber canines from the Permian of Brazil », Science, vol. 331, no 6024, , p. 1603-1605 (PMID 21436452, DOI 10.1126/science.1200305, Bibcode 2011Sci...331.1603C, S2CID 8178585, lire en ligne).
- ↑ (en) Kenneth D. Angielczyk et Christian F. Kammerer, « Non-Mammalian synapsids: the deep roots of the mammalian family tree », dans Frank E. Zachos & Robert J. Asher, Mammalian Evolution, Diversity and Systematics, Berlin, De Gruyter, , 392 p. (ISBN 978-3-110-27590-2), p. 151.
- ↑ a et b (en) Jörg Fröbisch et Robert R. Reisz, « The postcranial anatomy of Suminia getmanovi (Synapsida: Anomodontia), the earliest known arboreal tetrapod », Zoological Journal of the Linnean Society, vol. 162, no 3, , p. 661-698 (DOI 10.1111/j.1096-3642.2010.00685.x
 , S2CID 83780077).
, S2CID 83780077). - ↑ (en) Kenneth D. Angielczyk, « Phylogenetic evidence for and implications of a dual origin of propaliny in anomodont therapsids (Synapsida) », Paleobiology, vol. 30, no 2, , p. 268-296 (DOI 10.1666/0094-8373(2004)030<0268:PEFAIO>2.0.CO;2, S2CID 86147610).
- ↑ (en) Julien Benoit, Kenneth D. Angielczyk, Juri A. Miyamae, Paul Manger, Vincent Fernandez et Bruce Rubidge, « Evolution of facial innervation in anomodont therapsids (Synapsida): Insights from X-ray computerized microtomography », Journal of Morphology, vol. 279, no 5, , p. 673-701 (PMID 29464761, DOI 10.1002/jmor.20804, S2CID 3428692).
- ↑ a et b (en) Bruce S. Rubidge et James A. Hopson, « A primitive anomodont therapsid from the base of the Beaufort Group (Upper Permian) of South Africa », Zoological Journal of the Linnean Society, vol. 117, , p. 115-139 (DOI 10.1111/j.1096-3642.1996.tb02152.x , S2CID 85304536)
- ↑ a et b (en) Juan Carlos Cisneros, Fernando Abdala, Tea Jashashvili, Ana de Oliveira Bueno et Paula Dentzien-Dias, « Tiarajudens eccentricus and Anomocephalus africanus, two bizarre anomodonts (Synapsida, Therapsida) with dental occlusion from the Permian of Gondwana », Royal Society Open Science, vol. 2, no 7, , p. 150090 (PMID 26587266, PMCID 4632579, DOI 10.1098/rsos.150090 , Bibcode 2015RSOS....250090C)
- ↑ a b c et d (en) Kenneth D. Angielczyk et Christian F. Kammerer, « The cranial morphology, phylogenetic position and biogeography of the upper Permian dicynodont Compsodon helmoedi van Hoepen (Therapsida, Anomodontia) », Papers in Palaeontology, vol. 3, no 4, , p. 513-545 (DOI 10.1002/spp2.1087, S2CID 134092461)
- ↑ (en) Sean Modesto, Bruce Rubidge et Johann Welman, « The most basal anomodont therapsid and the primacy of Gondwana in the evolution of the anomodonts », Proceedings of the Royal Society of London B, vol. 266, no 1417, , p. 331-337 (PMCID 1689688, DOI 10.1098/rspb.1999.0642)

Voir aussi
Sur les autres projets Wikimedia :
- Chainosauria, sur Wikimedia Commons
- Chainosauria, sur Wikispecies
Articles connexes
Liens externes
- Ressource relative au vivant
 :
: - Paleobiology Database

 Portail de la paléontologie
Portail de la paléontologie