Bdelloidea
 Ten artykuł dotyczy wrotków potocznie zwanymi pijawczakami. Zobacz też: pijawczaki (rząd pijawek).
Ten artykuł dotyczy wrotków potocznie zwanymi pijawczakami. Zobacz też: pijawczaki (rząd pijawek).| Bdelloidea[1] | |||
| Hudson, 1884 | |||
 | |||
| Systematyka | |||
| Domena | eukarionty | ||
|---|---|---|---|
| Królestwo | zwierzęta | ||
| Podkrólestwo | |||
| Infrakrólestwo | pierwouste | ||
| Nadtyp | Spiralia | ||
| (bez rangi) | Gnathifera | ||
| (bez rangi) | Syndermata | ||
| Typ | wrotki | ||
| Gromada | Eurotatoria | ||
| Podgromada | Bdelloidea | ||
| |||
| |||
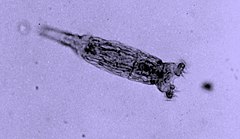
Bdelloidea – takson w randze gromady lub podgromady w typie wrotków (Rotifera). Znanych jest ponad 450 gatunków. 90% przedstawicieli typu Rotifera znajdywanych w glebie przynależy właśnie do Bdelloidea; oprócz tego żyją w liściach roślin mięsożernych, wśród bentosu, peryfitonu, ogółem w warstwie litoralu, biofilmu, w żwirowatych strumieniach, w mchu.
Osobniki żyjące w wodzie są przezroczyste, a te żyjące w mchu mogą być żółte, żółtobrązowe, czerwone, brązowe; jest to spowodowane występowaniem karotenów w ich organizmie. Potrafią zapaść w stan anabiozy, podobnie jak czynią to niesporczaki i nicienie, dzięki czemu mogą przetrwać okres braku pożywienia albo wody. Wrotki z tej podgromady tworzą wir za pomocą rzęsek, do którego spływają cząsteczki pokarmu, takiego jak jednokomórkowe glony lub bakterie, również grzyby, a jeden gatunek zjada inne wrotki, w tym innych przedstawicieli tej samej podgromady. Wrotki Bdelloidea rozmnażają się wyłącznie bezpłciowo i nie obserwowano, zarówno u populacji naturalnie występujących, jak i tych hodowanych w laboratoriach, samców. Bdelloidea mają sparowane gonady i rozmnażają się przez obligatoryjną partenogenezę; samice składają jaja, z których wylęgają się kolejne diploidalne samice.
W języku polskim określane są mianem pijawczaków.
Systematyka i filogenetyka

Takson został po raz pierwszy opisany przez mikrobiologa Charlesa Thomasa Hudsona w 1884 roku[2]. Swą nazwę wziął od pijawek (termin łaciński Bdella), które wrotki te przypominają[3][4]. Identyczną etymologię ma stosowana w języku polskim nazwa pijawczaki[5].
Początkowo na podstawie anatomii oraz sposobu poruszania się grupa ta została wyróżniona jako osobna gromada. W części źródeł Bdelloidea uznawana jest za podgromadę usytuowaną w gromadzie Eurotatoria oraz typie wrotków (Rotifera), inni podnoszą jednostkę do rangi gromady[6], jeszcze inni wyróżniają powyższy typ oraz gromadę, ale Bdelloidea łącznie z Monogononta degradują do rangi rzędu[7]. Podział taki proponował Voigt w 1957 roku i ostał się on do lat 90. XX wieku[3]. Następnie Markevich w roku 1993 utworzył podgromadę Archeorotatoria, do której zaliczył rząd Bdelloidea; jak argumentował, takson ten obejmuje formy prymitywne, które zachowały wiele cech podobnych do tych spotykanych u innych wrotków, ale ich układ wydalniczy jest zbliżony do tego spotykanego u płazińców (Platyhelmintes)[3]. W 1998 roku ukuto nazwę Platyzoa, która skupia w swoim obrębie bezkręgowce przynależące właśnie do Platyhelmintes, a także Gastrotricha oraz Gnathifera[8] (a ta z kolei zawiera: kolcogłowy, wrotki i szczękogębe[9]). Nie jest jednak uważana już za ważną wedle ITIS, tożsama z nią jest Spiralia[10].
Filogeneza pozostała słabo zbadana w przypadku tych bezkręgowców[11]. Badania molekularne, morfologiczne i embriologiczne pokazują, że omawiana grupa bezkręgowców to takson siostrzany wobec kolcogłowów (Acanthocephala), tworząc klad Lemniscea, albo, wedle innych analiz, Bdelloidea są pokrewne Monogononta, tworząc klad Eurotatoria[11]. Wyniki badania z 2008 roku ukazały, że Rotifera z Acanthocephala tworzą klad Syndermata, przy czym Bdelloidea są bliżej spokrewnione z kolcogłowami niż z Monogononta. Sugeruje się, że Monogononta oraz Bdelloidea zajęły początkowo odrębne nisze ekologiczne i że taksony rozwijały się w tym samym tempie, jednakże morfologia Monogononta zmieniała się szybciej, podczas gdy Bdelloidea zachowały wiele prymitywnych cech[3].
Bdelloidea są mniej zróżnicowaną grupą niż Monogononta – wyróżnia się 4 rodziny w jej obrębie, w tej drugiej rodzin jest 25[3]. Ponadto w obrębie rodzajów i gatunków różnice w morfologii są słabiej widoczne niż ma to miejsce u Monogononta. Bdelloidea mają dwie gonady, a Monogononta tylko jedną. W 1998 roku wykazano monofiletyzm Philodinida i bliskie jego pokrewieństwo z Habrotrochidae, więc w obrębie tego rzędu wyróżnia się również tę rodzinę. Wyróżnia się około 461 gatunków Bdelloidea na świecie[3] zgrupowanych w 18 rodzajach[4], a systematyka grupy pozostaje niezmieniona od 80 lat[11]. Poniżej zaprezentowany jest podział systematyczny wrotków Bdelloidea[3][2][1][6][11]:
Rząd: Adinetida (aparat rzęskowy w formie pola położonego po brzusznej stronie ciała[4])
- Rodzina: Adinetidae Hudson et Goose, 1889 (Bryce, 1910).
Rząd: Philodinavida (aparat rzęskowy ograniczony do podstawowego pola rzęskowego[4])
- Rodzina: Philodinavidae Harring, 1913.
Rząd: Philodinida (aparat rzęskowy w formie orzęsionych dysków służących do odżywiania się i pływania[4])
- Rodzina: Philodinidae Bryce, 1910,
- Rodzina: Habrotrochidae Bryce, 1910.
„Ewolucyjny skandal”
Przez Maynarda Smitha Bdelloidea wielokrotnie są nazywane „ewolucyjnym skandalem”, gdyż nie wyewoluowała u nich płeć[4]. Jest ona najdłużej istniejącą, największą i najbardziej zróżnicowaną grupą wielokomórkowych organizmów, która przetrwała bez występowania u niej płci. Najbardziej nietypową i oczywistą cechą Bdelloidea jest partenogenetyczny rozród[4].
Około 10% genomu Bdelloidea zajmują geny o pochodzeniu innym niż zwierzęce, które dostały się do niego na drodze horyzontalnego transferu genów. Jest to poziom o rząd wielkości większy niż u innych zwierząt[12].
Genetyka
Adineta vaga ma 12 chromosomów o mniej więcej tej samej wielkości i jest ona tetraploidem[13].
Budowa

Ogólna charakterystyka
Długość ciała waha się od 0,077 mm do 1,5 mm[3], chociaż inne źródła podają, że Bdelloidea osiągają minimalnie od 160 do 500–600 μm[14]. W ich ciele znajduje się blisko 1000 jąder komórkowych, ale posiadają neurony, wnętrzności i układ rozrodczy[4].
Wrotki z tej podgromady określane są jako podobne do robaków (worm-like)[15], są podłużne[6]. Całe ciało jest podzielone na pseudosegmenty – zawiera ono od 15 do 18 pseudosegmentów, w tym głowa zbudowana jest z trzech, szyja też z trzech, tułów z sześciu, noga z od trzech do sześciu[3]. Powłoka ciała jest zbudowana z tkanki nabłonkowej zbudowanej z syncytium, a dodatkowo tenże nabłonek produkuje rodzaj oskórka (kutykula); oskórek jest delikatny, ale wytrzymały oraz odporny na czynniki zewnętrzne i u niektórych gatunków zamieszkujących suchsze środowiska jest on twardy i sztywny, zaś u tych wodnych pozostaje elastyczny[3]. Bdelloidea z suchszych terenów mogą mieć liczne ziarnistości na oskórku oraz wyrostki w formie kolców, włosów, nitek, płetw i ramion – u innych ciało może być obsiane grudkami ziemi lub detrytusu, co nadaje im czarną lub brunatną barwę[3]. Niektóre gatunki Habrotrocha wydzielają przez nabłonek substancję, która krzepnąc tworzy charakterystycznego kształtu schronienie, nazywane muszlą wydzielinową[3].
Gatunki występujące w wodzie są przezroczyste, ale ciało przedstawicieli żyjących w wysuszonym mchu ubarwione jest w kolorach od czerwonego do brązowego lub od żółtego do żółtobrązowego[3]; Milne w pracy poświęconej Henoceros falcatus wspomniał, że opisany przez niego wrotek jest modry[16]. Również ciało wysokogórskich, arktycznych i antarktycznych Bdelloidea jest zabarwione. Ubarwienie mchowych wrotków jest związane z występowaniem ziaren karotenów, które przebywają na stałe w ścianach żołądka, w mniejszym stopniu w gonadach i jamie ciała; samice potrzebują karotenu, gdyż jest on transportowany do jaj[3].
Są to zwierzęta wykazujące eutelię[17].
Głowa, układ nerwowy i narządy zmysłu
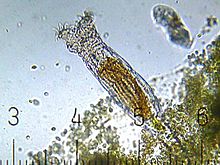
Głowa składa się z aparatu rzęskowego i rostrum[3].
Aparat rzęskowy (aparat wrotny zwany również koroną) jest cechą typową wrotków, występuje też u Bdelloidea; U rodzaju Embata aparat wrotny jest szerszy niż cały tułów, a u Habrotrochidae jest on zwykle mniejszy niż głowa i szyja, w przypadku pozostałych taksonów jest on proporcjonalny w stosunku do ciała lub nieco szerszy niż głowa[3]. Rzęski za pomocą swego ruchu mogą kierować cząsteczki pokarmu do otworu gębowego[3], ale aparat wrotny służy też do lokomocji[6]. Aparat rzęskowy składa się z: dwóch wieńców rzęsek (trochus), wargi dolnej z wieńcem rzęsek (cingulum), „kołnierza” posiadającego wybrzuszenie oraz z wargi górnej. Dwa wieńce rzęsek (zwane trochus) u wszystkich Bdelloidea rozdzielone są wgłębieniem (sulcus); może być ono różnej szerokości u poszczególnych rodzin. Czasami trochus posiada szczecinki czuciowe. Wieńce rzęsek znajdują się na szypułkach[3].
Mózg pozostaje skryty przez część przełyku; do układu nerwowego przydzielane również są też zwoje nerwowe gardzieli (mastaks), a także nogi oraz pnie nerwowe. U rodzaju Philodina zwój mózgowy przybiera kształt litery X[3].
Niektóre Bdelloidea posiadają o czerwonej (czerwień w różnych odcieniach) lub czarnej pigmentacji plamki oczne, zwane oczami, które można podzielić na: sparowane oczy mózgowe (leżące na zwoju mózgowym) i złożone z kilku par oczy rostralne (wierzchołkowe; położone wewnątrz rostrum). Mimo to, większa część wrotków Bdelloidea nie posiada plamek ocznych[3]. Taksony z tej podgromady mają też rzęski; można je podzielić na rzęski czuciowe na trochus, rostrum i rzęski na czułku grzbietowym – dodatkowo na rostrum występują dwie rzęski dotykowe[3].
Ruch drgający rzęsek aparatu wrotnego osiąga częstotliwość 8–12 Hz, w przeciwieństwie do Monogononta, u których ruch drgający rzęsek wynosi 14–27 Hz[3].

Reszta ciała
Szyja jest zbudowana z trzech podobnych budową pseudosegmentów, które są węższe od głowy[3]. Kilka gatunków rodzaju Habrotrocha mają na szyi poprzeczne bruzdy, a u jeszcze innych na brzusznej stronie pierwszego pseudosegmentu szyi występują wybrzuszenia tworzące pierścień; na nim umiejscowiony jest także czułek grzbietowy z budowany z 2–4 (rzadko) segmentów, który u poszczególnych gatunków przyjmuje inną wielkość oraz szerokość; na jego czubku znajdują się rzęski czuciowe. Natomiast w pozostałych segmentach szyi ulokowany jest mózg. W 3. pseudosegmencie znajduje się gardziel, która jest dobrze widoczna u wrotków posilających się[3].
Wszyscy przedstawiciele podgromady mają tułów złożony z 6 pseudosegmentów, pierwszy z nich jest najmniejszy, natomiast segmenty od 2 do 4 są najszersze, a ogółem ten typ segmentów pokryty jest mocniejszym oskórkiem, a u niektórych gatunków tworzy on swego rodzaju półpancerz[3]. Piąty i szósty pseudosegmenty tułowia są zwykle węższe niż trzy poprzednie. Czwarty segment przechodzi stopniowo w piąty lub to przejście jest bardzo gwałtowne i wtedy oba te segmenty są częścią nogi. Na krawędzi grzbietowej części szóstego segmentu usytuowany jest odbyt; 6. pseudosegment z 5. tworzą przyogonie, mogą też tworzyć biodro – razem z nogą mogą być wciągane do głównej części tułowia[3].
Noga posiada 2–4 palce, gruczoły czepne – są one umiejscowione na ostatnim pseudosegmencie. Do nogi przyczepione są również i ostrogi, które mogą być rozbudowane z innych pseudosegmentów, co tyczy się też palców – u rodzaju Rotaria ostrogi i palce są zbudowane z 2–3 segmentów. Ostrogi różnią się u każdego z gatunków wielkością, kształtem, odstępem między nimi; gatunki Habrotrocha acornis i Mniobia discophora są ich pozbawione[3]. Gruczoły czepne pozwalają przyczepiać się do podłoża[3].
Układ rozrodczy oraz wydalniczy
Układ wydalniczy składa się z 2–8 protonefrydiów, które z kolei złożone są z orzęsionych, dużych i płaskich komórek płomykowych. K. płomykowe uchodzą do kanalików, a te do pęcherza, który co jakiś czas wydala swoją zawartość do tylnej części kloaki[3].
Wrotki Bdelloidea mają 2 gonady, leżące blisko układu pokarmowego, po bokach ciała. Gonadę tworzy jajnik, gruczoł żółtkowy oraz warstwa folikularna – największą część gonady stanowi gruczoł żółtkowy, mający 8 jąder. Jajorodne wrotki mają rozwinięte jajowody, zaś w przypadku Rotaria rotatoria embriony rozwijają się w jamie ciała[3].
Występowanie
Tylko jeden gatunek zamieszkuje morza; najwięcej gatunków oraz rodzajów odnaleziono w Palearktyce[3].
Najsłabiej zbadanymi rejonami pozostają Meksyk, Tanzania oraz Australia[3], słabo zbadana grupa tych bezkręgowców występuje w Azji[14]. Odkrywane są też nowe gatunki w Europie.
W Polsce stwierdzono 85 gatunków z tej grupy, co stanowi niewielką część znanych z Europy[18].
Ekologia

Siedlisko
Są to ekstremofile[17]. Występują w środowiskach morskich, brachicznych, słodkowodnych i lądowych[2]; wyróżnia się zatem wrotki hydrofilne i kserofilne[3]. Znaleźć je można w słodkowodnych jeziorach, stawach, stwierdzono je też w strumieniach o żwirowatym dnie[16], w mchu, porostach, korze drzew, glebie i wśród detrytusu[4]. Ewolucyjnie, przodkowie tej podgromady wywodzili się z wody, ale opanowały one potem środowiska bardziej suche. 90% przedstawicieli wrotków znajdywanych w glebie przynależą do podgromady Bdelloidea[19]. Występują tutaj, gdy teren jest okresowo zalewany albo gdy jest wilgotno. Zwierzęta te także są spotykane w warstwie bentosu[20], albo wolno pływają w strefie litoralu – peryfitonu, pleustonu (wśród roślin z rodzaju Salvinia i Pistia) oraz heleoplanktonu, wśród glonów nitkowatych, biofilmu, psammonu[16], przyczepiają się też do kamieni lub muszli ślimaków[3]. Pawłowski (1984) wśród zgrupowań Lemna i Wolffia znalazł 140 gatunków wrotków, w tym 14 gatunków Bdelloidea[3]. Jeden z gatunków, Habrotrocha rosa jest znany z tego, że żyje w liściach Sarracenia purpurea[21].
Sposoby przetrwania
Wrotki jako ogół mogą tworzyć rodzaj cysty, w razie trudnych warunków lub w przypadku braku pożywienia[19]. Potrafią też formować schronienia ze swojego śluzu, do którego przylegają szczątki lub ekskrementy; inne mogą wykorzystywać opuszczone pancerzyki ameb skorupkowych[19]. Wrotkom z podgromady Bdelloidea przypisuje się „szereg niezwykłych cech”, spośród których wymienia się przede wszystkim: zdolność do horyzontalnego transferu genów oraz odporność na wyschnięcie, głód i promieniowanie jonizujące[3].
Wszystkie wrotki, w tym omawiany takson, potrafi zapaść, podobnie jak nicienie (Nematoda) i niesporczaki (Tardigrada), w stan anabiozy (anhydrobiozy), kiedy to pozbywają się wody z organizmu, mogą to czynić na każdym etapie swojego życia, w odpowiedzi na zmiany w środowisku[15]. Kiedy czynnik przyczyniający się do występowania anabiozy u wrotków zanika, te znowu nawadniają ciało i znów stają się aktywne[22][15]. Prawdopodobnie przodkowie Bdelloidea także potrafili zapaść w ten stan, jaja również mogą podlegać anhydrobiozie[3].
Badacze postawili hipotezę, że niestabilne warunki środowiska są nieodzowne, ażeby ich populacja przetrwała, ponieważ anhydrobioza jest warunkiem lepszego przeżycia. Z reguły u zwierząt i grzybów procesie anabiozy biorą udział polisacharydy, przykładowo trehaloza. U wrotków Bdelloidea nie stwierdzono jednak występowania trehalozy ani genu odpowiadającego za jej syntezę, a za anabiozę mogą odpowiadać białka LEA[3].
Odżywianie, ekologia i tryb życia

Wrotki te są konsumentami I rzędu[3]. Bdelloidea za pomocą rzęsek tworzą wir, do którego spływają cząsteczki pokarmu, tj. glony jednokomórkowe lub bakterie[15], także grzyby. Abrochtha carnivora, opisany naukowo w 2001 roku z Barbadosu jest drapieżcą[3] odżywiającym się innymi wrotkami (w tym innymi Bdelloidea), od czego zyskał swój epitet gatunkowy[23]. Swój pokarm w formie drobnych cząsteczek odfiltrowują, zeskrobują z podłoża lub aktywnie wyłapują[3]. W jednym z badań, dla kultur wrotków z gatunku Philodina roseola przeznaczono łatwo dostępny pokarm, glony z gatunku Raphidocelis subcapitata. Jeszcze w innym badaniu, dobrą pożywką dla Bdelloidea były bakterie, jak i drożdże, co zoptymalizowało wzrost badanych zwierząt. Jednak zauważono, że mimo że osobniki danego gatunku są klonem, a więc są identyczne genetycznie i morfologicznie, mogą mieć odrębne preferencje i wymagania[22].
Zwierzęta te atakowane są przez pleśnie, padają też ofiarą dużych ameb z taksonu korzenionóżek (Rhizopoda) i orzęsków (Ciliata)[3]. Pierwotniaki z gromady sysydlaczków (Suctoria) wysysają ciała martwych wrotków, zostawiając poszczególne części ciała po swojej uczcie, natomiast duże ameby aktywnie chwytają wrotki. Również i niesporczaki mogą zjeść młode okazy[3]. Wrotki te, pełzając po dnie, są trudnym obiektem do upolowania. Bdelloidea żyją też na innych organizmach, jako komensale[3]. Niektóre gatunki mają zdolność wytwarzania środków grzybobójczych o charakterze antybiotyków. Są one podobne do tyrocydyny A, ale na tyle odmienne, że przypuszczalnie geny je kodujące zostały przejęte od bakterii na tyle dawno, że zaszła dość duża ich ewolucja[12].
Potrafią się rozprzestrzeniać za pomocą wiatru, wody, jak i innych zwierząt[14][3]. Sposób poruszanie przypomina ten spotykany u pijawek[20] (Hirudinea); pełzają one lub łączą ten typ lokomocji z pływaniem, przy czym inaczej wygląda ten sam wrotek, gdy pożywia się, pływa lub odpoczywa[3]. Każdy wrotek z podgromady Bdelloidea potrafi pełzać, lecz nie każdy potrafi pływać[3]. Adinetidae używają swojej pozbawionej wieńców rzęsek korony tylko do zbierania pokarmu, ponieważ ruch rzęsek korony nie wywołuje u nich kierunkowego ruchu wody. Ciało odpoczywającego wrotka jest w kształcie kulki lub beczułki, ale przykładowo Rotaria neptunia podczas odpoczynku składa swe ciało jak teleskop[3].
Rozród
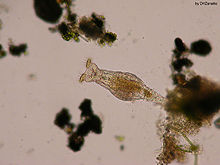
Badania rozrodu u Philodina roseola ujawniły, że organizm ten szybko dorasta. Ponadto, po pierwszym złożeniu jaj, osobnik macierzysty nie przestaje rosnąć, mimo że więcej energii przeznacza on na rozród niż na wzrost[22].
Już od dawna przypuszczano, że Bdelloidea są zwierzętami aseksualnymi. Tezę tę zasugerowano, gdyż nie obserwowano osobników płci męskiej zarówno u populacji żyjących naturalnie, jak i tych hodowanych w laboratoriach[24]. Dodatkowo, nie zaobserwowano u nich mejozy. Z kolei badania z 2022 roku zdają się sugerować, że zachodzi ona u tych bezkręgowców[17]. Badanie genetycznie przeprowadzone na Adineta vaga wykazało, że organizm ten jest tetraploidem[13]. Obecnie wiadomo, że wrotki Bdelloidea jako jedyni przedstawiciele Metazoa, rozmnażają się poprzez obligatoryjną partenogenezę[25]. Odmienne zdanie mają badania z 2022 roku, które kwestionują aseksualność tych wrotków, a reprodukcja płciowa może występować raz na jakiś czas, podobnie jak ma to miejsce u drożdży Saccharomyces cerevisiae, u których ta może występować raz na 25–50 000 generacji[17]. Przypuszcza się, że przodkowie Bdelloidea, podobnie jak przedstawiciele Monogononta, rozmnażali się głównie partenogenetycznie, ale co jakiś czas pojawiało się pokolenie rozmnażające płciowo, po czym utracili zdolność przemiany pokoleń[26]. Spotyka się u nich podobnie jak u morskich wrotków z gromady Pararotatoria, sparowane gonady[27]. Uważa się, że Bdelloidea przeszły rzeczywistą specjację mimo braku występowania rozmnażania płciowego[3]. U Bdelloidea nie występują mechanizmy usuwania mutacji oraz nie spotyka się u nich tworzenia nowych adaptacji do zmian środowiskowych, co jest jedną z „zalet” w przypadku występowania płci; mimo to, przetrwały miliony lat i z tego względu ewolucjoniści starają się badać środowisko i mechanizmy gatunkowe, aby poznać ich ewolucyjny sukces[28].
Karoten jest potrzebny u samic, gdyż jest on później transportowany do jaj[3]. Samice składają diploidalne jaja, przy czym komórki jajowe są produkowane raz przez jedną gonadę, czasem przez tę drugą[3]. Wielkość jaj waha się, nawet w obrębie gatunku – długość jaja Mniobia russeola waha się od 156,1 do 189 μm, w przypadku Habrotrocha constricta od 60,8 do 84,2, u Macrotrachela quadricornifera od 56 do 106,5[3], a u Henoceros falcatus 60[16]. Wewnętrzna cienka błona jaja pokrywa embrion; jaja są owalne w kształcie, o gładkiej powierzchni lub z wyrostkami, jednocześnie mogą być czasem spłaszczone po stronie brzusznej. Bruzdkowanie, podobnie jak jest to w przypadku innych wrotków, jest całkowite, przeprowadzane nierównomiernie, zbliżone do spiralnego i ściśle zdeterminowane[3]. W przypadku Habrotrocha rosa rozwój od złożenia jaja do wyklucia trwał 31,5 ± 1 godzin[3]. Małe zarodki potrzebują więcej czasu na rozwój w jaju niż ich większe odpowiedniki[3]. Z jaj wylęgają się diploidalne samice[3].
Philodina roseola w temperaturze 24 °C żyła 42 dni i złożyła 55 jaj, Philodina acuticornis składała średnio 52 jaja przy długości życia 16–22 dni i temperaturze 24 do 27 °C, a Habrotrocha rosa produkowała 30 jaj w ciągu od sześciu do dziewięciu dni; gatunki żyjące w wodzie krócej żyją, są bardziej płodne i szybciej dojrzewają niż te spotykane we mchach[3].
Zobacz też
Przypisy
- ↑ a b Bdelloidea, [w:] Integrated Taxonomic Information System [dostęp 2024-01-30] (ang.).
- ↑ a b c Bdelloidea, [w:] WoRMS – World Register of Marine Species [online] [dostęp 2024-01-25] (ang.).
- ↑ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am an ao ap aq ar as at au av aw ax ay az ba bb bc bd be IrenaI. Bielańska-Grajner IrenaI., JolantaJ. Ejsmont-Karabin JolantaJ., NataliiaN. Iakovenko NataliiaN., Wrotki (Rotifera, Bdelloidea): Zeszyt 32C, AndrzejA. Piechocki (red.), Katedra Zoologii Bezkręgowców i Hydrobiologii Uniwersytetu Łódzkiego 2013, DOI: 10.18778/7969-085-5, ISBN 978-83-7969-085-5, ISSN 0071-4089 (pol.).
- ↑ a b c d e f g h i C. Ricci & D.C.R.& D. Fontaneto C. Ricci & D.C.R.& D., The importance of being a bdelloid: Ecological and evolutionary consequences of dormancy, „Italian Journal of Zoology”, 2009, s. 240-249, DOI: 10.1080/11250000902773484 [dostęp 2024-08-06] (ang.).
- ↑ AnnaA. Hillbricht AnnaA., Dynamika populacji Philodina citrina Ehr. (Rotatoria) w hodowli akwariowej, Polska Akademia Nauk (PAN), KomitetK. Ekologiczny, Warszawa 1960, s. 161-170 [dostęp 2024-04-21], Cytat: Gatunek ten, podobnie jak większość Bdelloidea (pijawczaki), wykazuje dwa sposoby ruchu... (pol.).
- ↑ a b c d GiulioG. Melone GiulioG., ClaudiaC. Ricci ClaudiaC., Rotatory apparatus in Bdelloids, „Hydrobiologia”, 313, 1995, s. 91–98 [dostęp 2024-01-30] (ang.).
- ↑ JeremyJ. Wright JeremyJ., Rotifera – wheel or whirling animals (also: rotifers), [w:] ADW: Animal Diversity Web [online] [dostęp 2024-01-30] (ang.).
- ↑ Phylogeny of platyzoan taxa based on molecular data w: ThomasT. Hankeln ThomasT. i inni, Deep Metazoan Phylogeny, 2014, s. 105-125, DOI: 10.1515/9783110277524.105 [dostęp 2024-08-28] (ang.).
- ↑ Witeka A., Herlynb H., Ebersbergerc I., Mark Welchd D.B., Hankeln T. Support for the monophyletic origin of Gnathifera from phylogenomics. „Molecular Phylogenetics and Evolution”. 53(3), s. 1037-1041, 2009. DOI: 10.1016/j.ympev.2009.07.031. (ang.).
- ↑ Platyzoa, [w:] Integrated Taxonomic Information System [dostęp 2024-09-09] (ang.).
- ↑ a b c d EricaE. Lasek-Nesselquist EricaE., A Mitogenomic Re-Evaluation of the Bdelloid Phylogeny and Relationships among the Syndermata, „PLoS ONE”, 2012, DOI: 10.1371/journal.pone.0043554 (ang.).
- ↑ a b Reuben W.R.W. Nowell Reuben W.R.W. i inni, Bdelloid rotifers deploy horizontally acquired biosynthetic genes against a fungal pathogen, „Nature Communications”, 15 (1), 2024, DOI: 10.1038/s41467-024-49919-1, ISSN 2041-1723, PMID: 39025839, PMCID: PMC11258130 [dostęp 2024-08-10] (ang.).
- ↑ a b Eugene A.E.A. Gladyshev Eugene A.E.A., Irina R.I.R. Arkhipova Irina R.I.R., Genome Structure of Bdelloid Rotifers: Shaped by Asexuality or Desiccation?, wyd. 1, t. 101, 2010, s. 85–93, DOI: 10.1093/jhered/esq008 [dostęp 2024-01-28] (ang.).
- ↑ a b c YueY. Zeng YueY. i inni, Bdelloid rotifers (Rotifera, Bdelloidea) of China: diversity and new records, „ZooKeys”, 2020, DOI: 10.3897/zookeys.941.50465 [dostęp 2024-01-25] (ang.).
- ↑ a b c d David C.D.C. Coleman David C.D.C., Diana H.D.H. Wall Diana H.D.H., 7 - Fauna: The Engine For Microbial Activity And Transport, [w:] Soil Microbiology, Ecology and Biochemistry (Third Edition), s. 169, DOI: 10.1016/B978-0-08-047514-1.50011-1 [dostęp 2024-03-16] (ang.).
- ↑ a b c d J.M.J.M. Schmid-Araya J.M.J.M., New records of rare Bdelloidea and Monogononta rotifers in gravel streams, „Archiv fur Hydrobiologie”, 1, 135, Stuttgart 1995, s. 129–143 [dostęp 2024-01-25] (ang.).
- ↑ a b c d Veronika N.V.N. Laine Veronika N.V.N., Timothy B.T.B. Sackton Timothy B.T.B., MatthewM. Meselson MatthewM., Genomic signature of sexual reproduction in the bdelloid rotifer Macrotrachella quadricornifera, „Genomics”, 2, 220, 2022, DOI: 10.1093/genetics/iyab221, PMID: 34888647 [dostęp 2024-08-22] (ang.).
- ↑ Wykaz zwierząt Polski. Razowski J. (red.). T. IV. Kraków: Instytut Systematyki i Ewolucji Zwierząt PAN, 1997, s. 303. ISBN 978-83-907187-0-5.
- ↑ a b c David C.D.C. Coleman David C.D.C., D.A.D.A. Crossley D.A.D.A., Paul F.P.F. Hendrix Paul F.P.F., Secondary Production: Activities of Heterotrophic Organisms—The Soil Fauna, Elsevier, 2004, s. 79–185, DOI: 10.1016/b978-012179726-3/50005-8 [dostęp 2024-03-16] .
- ↑ a b R.L.R.L. Wallace R.L.R.L., H.A.H.A. Smith H.A.H.A., Rotifera, Gene E.G.E. Likens (red.), Oxford: Academic Press, s. 689–703, DOI: 10.1016/b978-012370626-3.00147-2, ISBN 978-0-12-370626-3 [dostęp 2024-03-16] .
- ↑ L.L. Bateman L.L., A bdelloid rotifer living as an inquiline in leaves of the pitcher plant, Sarracenia purpurea, „Hydrobiologia”, 147, 1987, s. 129–133, DOI: 10.1007/BF00025735 [dostęp 2024-01-25] (ang.).
- ↑ a b c Raquel A.R.A. Moreira Raquel A.R.A., Adrislaine S.A.S. Mansano Adrislaine S.A.S., OdeteO. Rocha OdeteO., Life cycle traits of Philodina roseola Ehrenberg, 1830 (Rotifera, Bdelloidea), a model organism for bioassays, „Biological Sciences”, 2016, DOI: 10.1590/0001-3765201620140729 [dostęp 2024-01-25] (ang.).
- ↑ ClaudiaC. Ricci ClaudiaC., GiulioG. Melone GiulioG., Elizabeth J.E.J. Walsh Elizabeth J.E.J., A Carnivorous Bdelloid Rotifer, Abrochtha carnivora n.sp., wyd. 2, t. 120, 2001, s. 136-141, JSTOR: 3227112 [dostęp 2024-01-28] (ang.).
- ↑ ManuelM. Serra ManuelM., Terry W.T.W. Snell Terry W.T.W., Robert L.R.L. Wallace Robert L.R.L., Reproduction, Overview by Phylogeny: Rotifera, Michael K.M.K. Skinner (red.), Oxford: Academic Press, s. 513–521, DOI: 10.1016/b978-0-12-809633-8.20646-8, ISBN 978-0-12-815145-7 [dostęp 2024-03-17] .
- ↑ ManuelaM. Pagani ManuelaM., Claudia Ricci & Carlo AlbertoC.R.& C.A. Redi Claudia Ricci & Carlo AlbertoC.R.& C.A., Oogenesis in Macrotrachela quadricornifera (Rotifera, Bdelloidea), „Rotifer Symposium VI (Part of the Developments in Hydrobiology; vol. 83)”, 1993, s. 225–230 [dostęp 2024-01-26] (ang.).
- ↑ C. WilliamC.W. Birky C. WilliamC.W., Bdelloid rotifers revisited, „Proceedings of the National Academy of Sciences of the United States of America”, 101 (9), 2004, s. 2651–2652, DOI: 10.1073/pnas.0308453101, ISSN 0027-8424, PMID: 14981265, PMCID: PMC365675 [dostęp 2024-02-21] (ang.).
- ↑ ClaudiaC. Ricci ClaudiaC., GiulioG. Melone GiulioG., CristinaC. Sotgia CristinaC., Old and new data on Seisonidea (Rotifera), „Hydrobiologia”, 1993, s. 495-511, DOI: 10.1007/BF00025879 [dostęp 2024-08-20] (ang.).
- ↑ ManuelM. Serra ManuelM., Terry W.T.W. Snell Terry W.T.W., Robert L.R.L. Wallace Robert L.R.L., Reproduction, Overview by Phylogeny: Rotifera, t. 6, 2018, s. 513-521, DOI: 10.1016/B978-0-12-809633-8.20646-8 [dostęp 2024-08-05] (ang.).
- Britannica: animal/Bdelloidea








